AJAA Index | Virtual Library | Magazine Rack | Search
Abstract. Microbial activities important to effects on crop productivity and nutrient cycling can be altered by agricultural management practices. This study was conducted to determine whether soil microbial populations and their N cycling activities differ between conventional and alternative management practices. Physical, chemical, and microbial soil properties were measured at soil depth intervals of 0 to 7.5, 7.5 to 15, and 15 to 30 cm at a site in southeastern Pennsylvania during the second and fifth years after conversion from a conventional, chemically intensive system to alternative systems utilizing legumes and animal manure as N sources. In the second year after conversion, populations of fungi and bacteria, dehydrogenate activity, and soil respiration in the surface soil layer were greatest with alternative systems planted to red clover (Trifolium pratense L.). Differences in soil biological factors between management systems were related primarily to crop characteristics and, to a lesser extent, to soil physical properties. Levels of microbial populations and activities with conventional management were the same as with alternative management systems when similar crops such as corn (Zea mays L.) or soybean [Glycine max (L.) Merrilll were grown. Soil NO3-N contents, at most sampling depths, were markedly increased by application of fertilizer N or recent plow-down of red clover or hairy vetch (Vicia villosa Roth). The growth of red clover in the second year or hairy vetch in the fifth year was accompanied by significantly increased microbial biomass and potentially mineralizable N (PMN) reserves in the top 30-cm soil layer--these changes being most pronounced in the surface 0- to 7.5-cm layer. Nitrogen deficiency symptoms and longer corn grain yields in a legume/cash grain rotation as compared with conventional management in the second year were associated with longer soil NO3 levers and a greater proportion of N present as weed biomass and below ground microbial biomass. In 1985, management systems comparisons were limited to corn as the main crop; soil NO3 levels during the growing season were inversely related to soil microbial biomass and PMN levels where hairy vetch was overseeded and incorporated as green manure by plowing before corn planting. Under the conditions of this study, the use of chemicals had little effect on microbial populations, their activity, or the cycling of nitrogen. Cropping systems--in particular, the growth of red clover or hairy vetch--profoundly influenced soil microbial biomass levels and soil pools of organic and available NO3-N during the growing season. Competitiveness of alternative management systems employing legumes as N sources for grain crops may depend largely on the grower's ability to synchronize supplies of available soil N with periods of maximum uptake by grain crops.
The productivity and stability of soil as a medium for plant growth depend greatly on the balance between living and nonliving components. Energy from the sun and nutrients essential for growth, stored in the fabric of crop plants, are recovered for reuse through decomposition activities of micro- and macroorganisms in soil. The soil organic master formed during this process serves both as a continuous nutrient supply and a factor stabilizing the soil physical environment. To maintain productivity, soluble nutrients removed from soil through plant growth and harvest must be replaced, either as fertilizers or through biological decomposition of organic and plant master in the soil. In management systems in which synthetic chemical use is reduced or eliminated, it follows that the action of soil microorganisms and fauna becomes a major determinant of nutrient cycling and plant growth.
Advocates of alternative or organic agricultural management emphasize the importance of on-farm recycling of nutrients for crop production and the critical effect of proper balance between organic master, soil organisms, and plant diversity on productivity of soils. A basic tenet of organic farming is that an ecologically balanced soil environment results in healthy, vigorous plants and a stable soil environment (U.S. Dept. of Agriculture, 1980). Proponents of this system think the use of synthetic chemical fertilizers and pesticides is harmful to microorganisms and other soil life forms and such use is therefore avoided. In experimental tests, however, the use of chemical fertilizers or pesticides, at recommended application rates, has shown little direct long-term effect on microbial populations and their activity in nutrient cycling (Goring and Laskowski, 1982; U.S. Dept. of Agriculture, 1980). Decreased soil productivity associated with cash crop monoculture, a management system often employing intensive use of chemical fertilizers and pesticides, more probably results directly from increased erosion, reduced plant cover, increased disease incidence, and greater export of nutrients in response to cultural management practices.
Alternative agricultural cropping systems are being sought which will reduce on-farm inputs to offset rising production costs, decrease environmental and health hazards associated with use of agricultural chemicals, and maintain soil fertility and productivity levels. Management systems using animal residues and legume and green-manure crops in rotation, as alternatives to synthetic chemicals, may require larger farm acreage to achieve cash grain yields comparable to those with chemically intensive management (Council for Agricultural Science and Technology, 1980). Organic farming practices conserve C and N in the soil-plant system, and reduced net mineralization may result in mild N stress during periods of rapid plant growth (Power and Doran, 1984). Change from conventional to organic management systems may require a transition period during which the soil ecosystem adjusts to a new equilibrium, before comparable yields are achieved (Culik, 1983). Limitations on crop yields during transition from conventional to alternative management include: (i) reduced N availability; (ii) increased weed infestations; and (iii) incomplete user adjustment to new management techniques (Regenerative Agriculture Association, 1985; U.S. Dept. of Agriculture, 1980).
The research reported here was part of an interdisciplinary research project initiated by the Rodale Research Center in 1981 to study yield-limiting factors faced by farmers during transition from conventional to alternative organic management cropping systems. A major research objective was to characterize biological changes occurring during the transition phase from conventional to alternative practices. Microbial populations and their activities in relation to N cycling in soil were the variables studied. A secondary objective was to determine whether use of chemical fertilizers and pesticides directly reduced microbial populations and activities in the field.
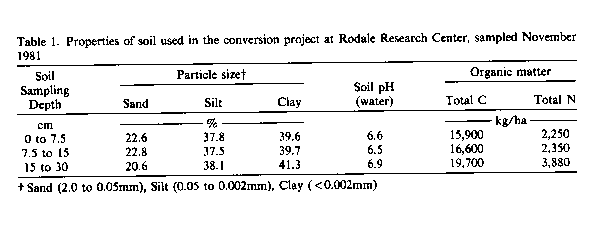
The research study site was at the Rodale Research Center at Kutztown in southeastern Pennsylvania. The overall project, known as "the conversion project", was initiated in 1981 on land which previously had been farmed for production of corn and wheat (Triticum aestivum L.) by the use of conventional management practices. The climate of the area is humid, continental with a mean annual temperature of 12.4°C and a mean annual precipitation of 108 cm, of which 46 percent falls between May and September. The soil at the experimental site is predominantly a Comly silt loam (Typic Fragiudalf) with less than 5 percent slope. Physical and chemical characteristics of surface soil (30 cm) are shown in Table 1.
Three different farming systems were compared in the conversion project (Table 2). The first, grain/forage rotation plus manure, in which N and other nutrients were replenished by the use of animal manure and legumes in rotation, simulated an organic operation in which field crops are raised mainly for beef and dairy cattle. The second system, legume/ cash grain rotation, simulated an organic cash grain farm without manure in which the only supplemental N was from plow-down of legumes. The third system, conventional cash grain, used synthetic chemical fertilizers to supply nutrients and herbicides to control weeds in a corn/soybean rotation. Each treatment consisted of a 5-year crop rotation split for three entry points into the cropping sequence to determine the viability of a rotation as a function of the crop with which it started. Further details on experimental design and management for the conversion project are provided by Culik (1983) and Regenerative Agriculture Association (1985).

Soil samples for microbial, chemical, and physical characterization were collected on November 10-12, 1981, June 24-26 and July 27-29, 1982, and four dates between April 10 and July 19, 1985 from four of eight replicates of the conversion project. Each sample represented a composite of 20 cores (2-cm diameter) taken at depths of 0 to 7.5, 7.5 to 15, and 15 to 30 cm. Samples were sealed in plastic bags and transported by air freight to the U.S. Department of Agriculture laboratory at Lincoln, Nebraska, where samples were stored at 22°C, and biological assays were begun within 2 days of sampling. Soil analyses were conducted on samples which had passed a 2 - mm sieve . Determinations of microbial biomass, microbial courts, and dehydrogenase activity were begun the same day as sieving. Soil was air dried for determination of nitrate-N (NO3-N) and ammonium-N (NH4-N) levers, soil pH, soluble P. particle size analysis, total organic C, and total Kjeldahl N within 1 day of sieving and stored until analyses could be conducted. Air dried soil samples used for determination of total C and N were "round to pass a 0.5-mm sieve before analysis. Details of chemical analysis procedures are provided by Doran (1980).
Soil microbial biomass was determined by the chloroform fumigation incubation method of Jenkinson and Powlson (1976). Potentially mineralizable nitrogen (PMN) was estimated from the ammonium released from soil during 16 hours of autoclaving at 121°C (Doran, 1987). Populations of fungi and bacteria were determined by dilution plate counting as described by Doran (1980) and soil dehydrogenase activity by the method of Casida et al. (1964). In-field measurements of soil respiration were made by gas chromatographic determinations of carbon dioxide accumulating in the headspace of 15-cm diameter cylindrical gas reservoirs, inserted 10 cm into the soil, over a 24hour period. Additional procedural details are given by Fraser (D. G. Fraser, M. S. thesis, University of Nebraska-Lincoln, 1984).
Results and discussion
Management comparisons among all crops and rotation entry points for physical, chemical, and biological soil properties were restricted to 1981 and 1982 during the early transition period from conventional to alternative management. Initial interpretations of these data indicated that crop type and growth characteristics complicated management comparisons. Consequently, comparisons made in 1985 were restricted to management treatments planted to corn, and sampling times during the growing season were increased to four. Discussion of results will emphasize comparisons during the early transition period (1981 and 1982), and discussion of the 1985 cropping season will be included as needed for clarification of earlier findings. A more detailed discussion of 1985 comparisons will appear elsewhere.
Soil properties in 1981 and 1982 varied with time of year, depth .n soil, and farming management system. Soil bulk density did not vary with time, but ranged from 0.98 to 1.17 and 1.14 to 1.27 Mg/m3 for the 0- to 15-cm and 15to 30-cm soil depths, respectively. Lower bulk densities near the soil surface (p < 0.05) were associated with crop rotation systems planted to red clover or those in conventional management. Volumetric soil water contents across dates and sampling depth ranged from 25.7 to 36.5 percent. Significant differences between management systems occurred near the soil surface (0 to 7.5 cm), where water contents of systems with legumes in rotation were 1.5 to 3.0 percent higher than those in conventionally managed soils. Soil pH, as measured in 0.01 M CaCl2, ranged from 5.6 to 6.0 across depths and management systems. Soil pH in the conventional systems with either corn or soybean was 0.1 to 0.3 pH units longer than with organic rotations in 1981 and 1982; lowest pH values occurred where ammoniacal fertilizer had been applied recently to corn.
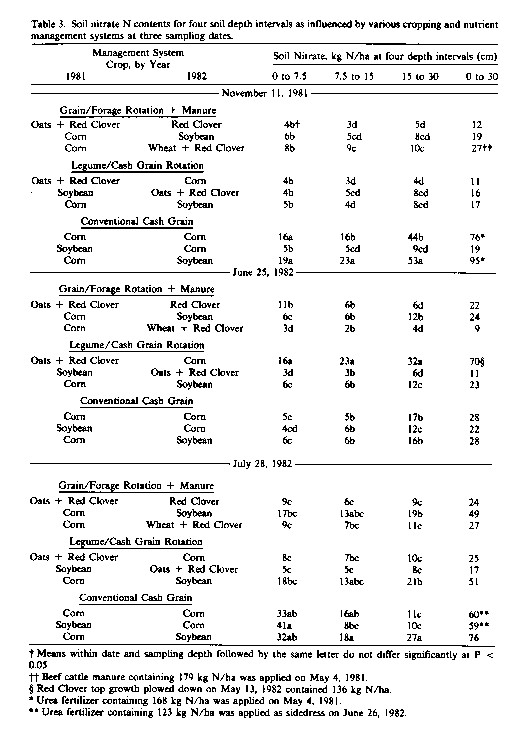
Significant differences in soil levels of plant-available nutrients between management systems in 1981 and 1982 were limited primarily to nitrate-N (NO3-N). Soluble plant-available forms of P and K in the surface 30 cm of soil ranged from 334 to 436 and 497 to 942 kg/ha, respectively, well within the range sufficient for crop growth. Soil ammonium N (NH4-N) levers did not vary significantly between management systems and averaged 37 and 41 kg N/ha in the top 30 cm of soil in 1981 and 1982, respectively. Soil NO3-N levels at most depths sampled, however, were markedly increased by application of fertilize' N or recent plow-down of clover (Table 3). Soil NO3-N levels at the 7.5- to 15 and 15- to 30-cm sampling depths in fall 1981 were also slightly greater in the treatment receiving beef cattle manure in the previous spring. During the 1982 growing season, corn in organic rotations not receiving animal manure or legume plow-down was noticeably yellow and, presumably, N-deficient. Soil NO3 levers, as summed across the 0- to 30-cm layers of soil for July 1982, were highly correlated (r2 = 0.88, p < 0.001) with the N content of corn leaves at silking as determined by the cooperating researchers (Regenerative Agriculture Association, 1985). These results seem to confirm earlier observations that corn growth can be limited by N during transition to organic management where supplemental N is not added.
The major organic pools of C and N in soil were apparently little influenced by management practice or cropping.
Levels of C and N. as given in Table 1, are representative of all management systems and cropping and varied little between samples taken in 1981, 1982, or 1985. Levels of Potentially Mineralizable Nitrogen (PMN) in the surface 0- to 7.5-cm soil layer averaged from 292 to 393 kg N/ha in 1981 and 1982 (data not shown). There was no significant difference between farming management systems for either 1981 or 1982, but there was a significant difference (p < 0.05) between crops in July 1982. Soils planted to red clover contained up to 80 kg PMN/ha more PMN than those planted to soybean. Highest levers of mineralizable N coincided with time of year when crop roots should have attained the greatest mass.
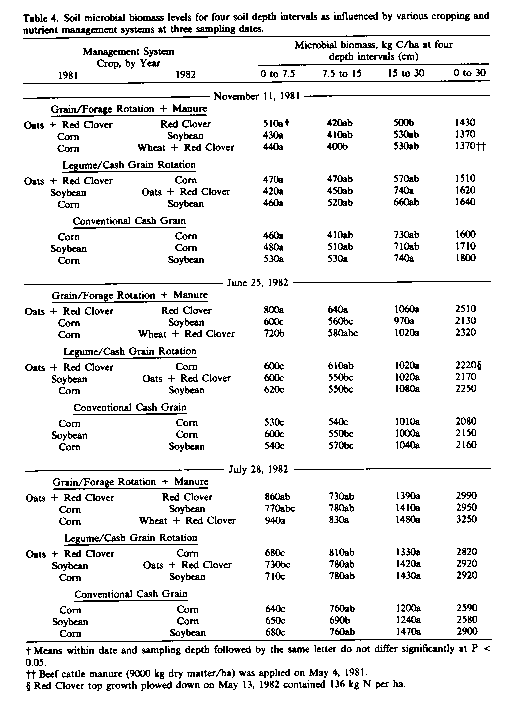
Differences in soil microbial biomass levers with varying nutrient management systems were observed primarily in 1942 in the surface 0- to 7.5-cm soil layer and, to a lesser extent, at 7.5 to 15 cm (Table 4). Microbial biomass levels were generally greatest in treatments planted to red clover and least in those planted to corn or soybean regardless of nutrient management system. The exception of longer biomass levels in the legume/cash grain rotation planted to oats and red clover in June 1982 may have resulted, at least in part, from a less optimal soil water statue for microbial activity as will be discussed later.
Microbial biomass levels, averaged across sampling depths and treatments, differed significantly between sampling dates (p < 0.05) and increased 464 and 984 kg C/ha, respectively, between postharvest in 1981 and June and July 1982. These increases in biomass C, which represent a potential sink for up to 115 kg/ ha of N. were likely related to increased plant activity between these time periods. Researchers in Canada and England have demonstrated a direct relationship between soil microbial biomass levers and plant growth and rooting activity (Carter and Rennie, 1984; Lynch and Panting, 1980).

The overall ecological productivity of an area of land is the sum of total biological growth over an interval of time. A common measure of agricultural productivity is grain yield of the crop plant. In 1982, corn grain yields from the conventional cash grain system of the conversion project, averaged 9,070 kg/ha compared with 5,470 kg/ha in the legume/cash grain rotation--an apparent 40 percent decline in productivity. Weed competition was identified as the major factor for decreased yields with the organic system. Measurement and estimates of total above- and belowground yields of corn, weeds, and microbial biomass would suggest, however, that noncrop biomass was also a significant pool of N (Table 5). Weeds and microbial biomass in the legume/cash grain rotation contained 49 and 38 kg/ha more N. respectively, than in the conventional management system. Difference in total N yields (above- and belowground) between the two systems was only 28 kg N/ha--about what might be expected from the additional input of fertilizer N with conventional management. Consequently, differences in total biological productivity between the two management systems may be far less than was indicated by differences in aboveground grain yields.
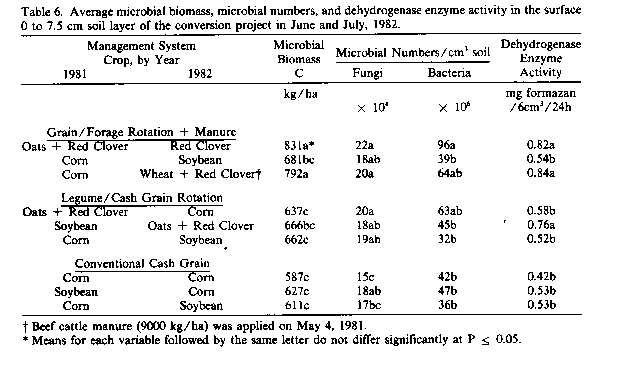
In 1982, characterization of microbial populations and activities were restricted to the surface 0- to 7.5-cm soil layer because earlier observations of microbial biomass indicated this to be the layer most influenced by management. Although numbers and activities were slightly longer in June than July of 1982, neither these differences nor interactions between dates and treatments were significantly different. For simplicity, the data given in Table 6 represent averages across sampling dates. Microbial biomass, fungal and bacterial courts, and dehydrogenase activities were generally highest in soils planted to red clover and least in those planted to corn or soybean. Thus, the type of crop grown appeared to have a greater influence on microbial numbers and activity during the cropping season than either the source of fertilizer N or the use of synthetic chemicals per se. Microbial numbers and activity were lowest in soils cropped to corn or soybean regardless of management system.
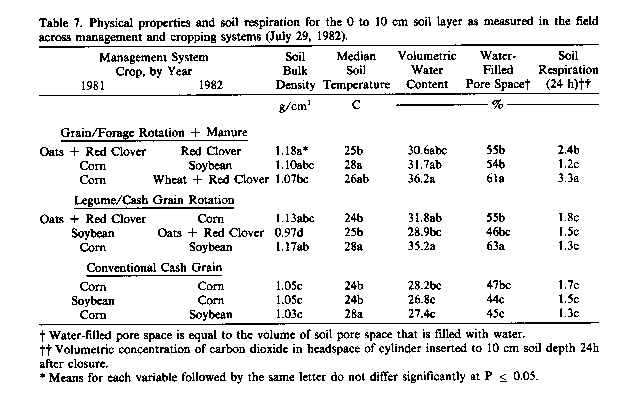
Measurement of soil respiration in the field enabled evaluation of the effect on microbial activity of management-related changes in the soil physical environment (Table 7). As occurred with microbial populations and soil dehydrogenase activity, respiration was highest in soils planted to red clover, with little difference between soils planted to corn or soybean regardless of management system. The highest respiration occurred from the soil planted to wheat and red clover, which had received 9,000 kg/ha of beef cattle manure the year before.
It is difficult to identify major factors controlling soil respiration because, in addition to the effects of soil environ -mental factors such as soil density, water statue, temperature, and organic substrates, it is influenced to varying extent by the activity of both soil microorganisms and plant roots. Activity of both microorganisms and plant roots increases with increasing temperature between 20° and 35°C and both achieve a maximum activity when about 60 percent of the soil pore space is water-filled-the point at which availability of water and air is maximal (Linn and Doran, 1984). Thus, the unusually low soil respiration from the soil planted to oats and red clover probably was a consequence of a longer soil density and water content, which resulted in a water-filled pore space comparable to that in soils planted to corn and soybean. The variations in microbial biomass or dehydrogenase were most likely representative of differences in carbonaceous substrates sup plied by crop plant roots, which also contributed to soil respiration. The higher respiration obtained with wheat and red clover may have resulted from more optimal soil water statue and supply of substrates from manure applied the previous year. Denitrification losses from intact cores, incubated at constant temperature in the laboratory, were also greatest from soil planted to wheat and red clover that had received manure, (data not shown).
Shifts in microbial numbers and activity in soil are often related to change in carbon inputs to soil as a result 0 management-related changes in crop type or residue addition. Soil microbial populations are increased by addition o animal manures or by synthetic fertilizers where crop production is simultaneously increased (Martyniuk and Wagner, 1978). Farming management systems which increase belowground in puts of C and N through inclusion o legumes and/or fibrous-rooted crops rotation sequence often increase microbial populations and activity above. that observed for conventional management with commercial fertilizers (Bolton et al., 1985).
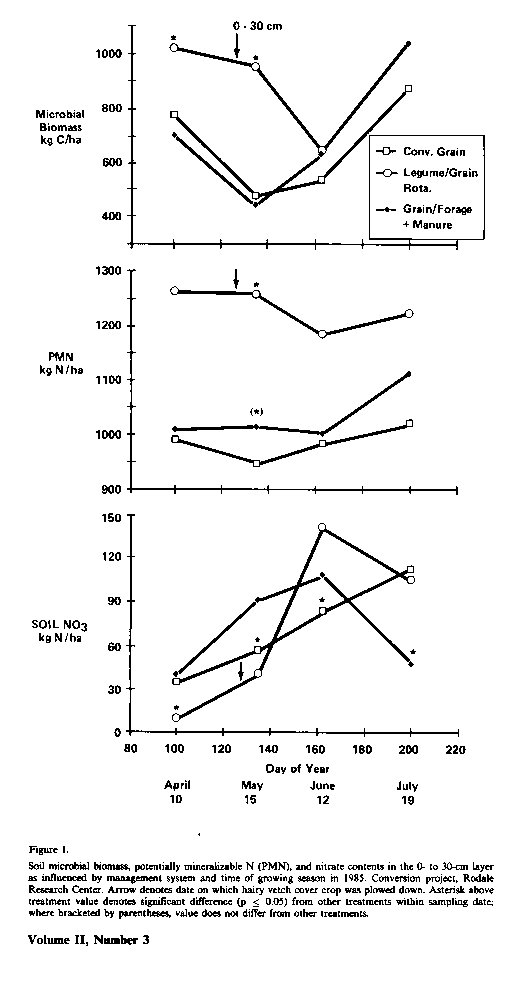
Biochemical comparisons between farming management systems in 1984 were limited to soils planted to corn to permit evaluation of effects of management without the confounding influence of type of crop grown. For the legume cash grain rotation systems, however,; hairy vetch cover crop, which had been seeded on September 6, 1984, was plowed under on May 8, 1985, 1 week before corn planting. In 1985, microbial biomass, PMN, and NO3-N levels in soil were significantly (p < 0.05) influence' by management practice and time of season (Fig. 1). Soil microbial biomass levels decreased as the season progressed until after June 10, at which time the increased to preseason levels or higher Fluctuations in microbial biomass level during the growing season were paralleled by changes in the opposite direction in NO3-N. The markedly greater quantity of soil biomass for corn with hairy vetch (281 kg C/ha) represents an additional pool of immobilized organic N. which, assuming a microbial biomass C/N of 8.5/1, is equivalent to 33 kg N/ ha. Preseason soil NO3-N levers for the legume/cash grain rotation system were 33 to 43 kg N/ha longer than with the grain/forage rotation with manure or the conventional management practice. Organic reserves of PMN for corn with hairy vetch averaged 222 kg N/ha greater than the other systems.
Early season soil NO3-N levers (April 10 and May 15), before and at planting, for the legume/cash grain rotation may well have been limiting for maximum growth of corn. The large increase in NO3-N (103 kg N/ha) after plow-down of the cover crop apparently resulted from mineralization of organic N. Part of this mineralized N probably came directly from microbial biomass, which decreased an equivalent of 37 kg N/ha during the same time period. Declines in soil NO3-N for the organic management practices between June and July amounted to 43 to 73 kg N/ha and were accompanied by increases in microbial biomass equivalent to 46 to 48 kg N/ ha. The relatively high soil nitrate-N levels in the conventional cash grain system is likely a reflection of the 112 kg of NH4 NO3 fertilizer N. which was applied as a side dress on June 17.
One interpretation of these data supports the importance of understanding the microbial immobilization/mineralization phenomenon in cycling of N and control of N availability to cash grain crop plants. Inclusion of legumes in a cropping system can dramatically influence the quantity and quality of soil microbial biomass and organic master (Biederbeck et al., 1984). Also, as demonstrated by McGill et al. (1986), seasonal fluctuations in the soil physical environment may be closely related to fluctuations in microbial biomass and mineralization of organic N pools in soil. The production of cash grain crops, in management systems where plant-available N is not supplemented with commercial fertilizers, may be regulated by type of crop grown and fluctuations in the soil physical environment during the growing season.
Microbial populations and activities in surface soil of alternative and conventional management systems were primarily influenced by type of crop grown and, to a lesser extent, by soil physical conditions. Under the conditions of our study, there was little observable detriment to microbial activity from use of chemicals per se. Soil microbial biomass levers and PMN reserves were greatest in alternative management systems planted to red clover or hairy vetch. Nitrogen deficiency symptoms and longer corn grain yields in the legume/cash grain rotation, as compared with conventional management during the early conversion period, were associated with longer soil NO3 levers and a greater proportion of N present in weed biomass and belowground microbial biomass pools. The immobilization/mineralization phenomenon catalyzed by soil microorganisms is important to recycling of soil organic N and subsequent availability to grain crops. The productivity of alternative management systems employing legumes as sources of N may depend largely on the grower's ability to synchronize supplies of available soil N with periods of maximum uptake by grain crops. In this regard, management of timing and type of tillage for soil incorporation of legume cover crops appears to be an important determinant of grain production efficiency in alternative management systems.
Acknowledgment. This paper is a contribution from the Agricultural Research Service, U. S. Department of Agriculture, in cooperation with the Nebraska Agricultural Research Division, University of Nebraska-Lincoln, Lincoln, Nebraska. Published as Journal Sciences No. 8369. Appreciation is expressed to J. K. Radke and the staff at Rodale Research Center for their foresight and vigilance in establishing and maintaining farming management comparisons. A special thanks is extended to Brian Bahler, Andrew Olson, and Jim McManus for capable technical assistance, which assured the successful completion of this project.
1. Biederbeck, V. O., C. A. Campbell, and R. P. Zenter. 1984. Effect of crop rotation and fertilization on some biological properties of a loam in southwestern Saskatchewan. Can. J. Soil Sci. 64:335-367.
2. Bolton, H., Jr., L. F. Elliott, R. I. Papendick, and D. F. Bezdicek. 1985. Soil microbial biomass and selected soil enzyme activities: Effect of fertilization and cropping practices. Soil Biol. Biochem. 17:297302.
3. Carter, M. R., and D. A. Rennie. 1984. Dynamics of soil microbial biomass N under zero and shallow tillage for spring wheat, using N urea. Plant Soil 76:157-164.
4. Casida, L. E., Jr., D. A. Klein, and T. Santoro. 1964. Soil dehydrogenase activity. Soil Sci. 98:371-376.
5. Council for Agricultural Science and Technology. 1980. Organic and conventional farming compared. Report No. 84. Council for Agricultural Science and Technology, Ames, Iowa. 32 pp.
6. Culik, M. N. 1983. The conversion experiment: Reducing farming costs. J. Soil Water Conserv. 38:333
7. Doran, J. W. 1980. Soil microbial and biochemical changes associated with reduced tillage. Soil Sci. Soc. Am. J. 44:765-771.
8. Doran, J. W. 1987. Microbial biomass and mineralizable nitrogen distributions in no-tillage and plowed soils. Biol. Fertility Soils 5:68-75.
9. Goring, C. A. I., and D. A. Laskowski. 1982. The effects of pesticides on nitrogen transformations in soils. In F. J. Stevenson (ed.) Nitrogen in agricultural soils. Agron. Monograph 22. Am. Soc. Agron., Madison, Wisconsin. pp. 689-720.
10. Jenkinson, D. S., and D. S. Powlson. 1976. The effect of biocidal treatments on metabolism in soil. V. A. method for measuring soil biomass. Soil Biol. Biochem. 8:209-213.
11. Linn, D. M., and J. W. Doran. 1984. Effect of water filled pore space on CO2 and N:O production in tilled and nontilled soils. Soil Sci. Soc. Am. J. 48:12671272.
12. Lynch, J. M., and L. M. Panting. 1980. Cultivation and the soil biomass. Soil Biol. Biochem. 12:29-33.
13. Martyniuk, S., and G. M. Wagner. 1978. Quantitative and qualitative examination of soil microflora associated with different management systems. Soil Sci.
14. McGill, W. B. K. R. Cannon, J. A. Robertson, an F. D. Cook. 19;36. Dynamics of soil microbial biomass and water-soluble organic C in Breton L after 50 years of cropping to two rotations. Can. J. Soil Sci. 66:1 19.
15. Power, J. F., and J. W. Doran. 1984. Nitrogen us in organic farming. In R. D. Hauck (ed.) Nitrogen in crop production. Am. Soc. Agron., Madison, Wisconsin. Chap. 40, pp. 585-598.
16. Regenerative Agriculture Association. 1985. The dollars and sense of resource-efficient farming. In Brusko (ed.) Profitable farming now! Regenerative- Agric. Assoc., Emmaus, Pennsylvania. pp. 29-45.
17. U.S. Dept. of Agriculture. 1980. Report and recommendations on organic farming. U.S. Govt. Printing Office, Washington, DC. 94 pp.
Citation : Doran John W., Fraser Daniel F., Culik Martin N., Liebhardt William C., 1987 " Influence of alternative and conventional agricultural management on soil microbial processes and nitrogen availability". Vol 2, No. 3, pp. 99-106.
Copyright © 1987 Reprinted with permission.
Reprinted with permission.
Info Request | Services | Become EAP Member | Site Map
Give us your comments about the EAP site
Ecological Agriculture Projects, McGill University (Macdonald
Campus)
Ste-Anne-de-Bellevue, QC, H9X 3V9 Canada
Telephone:
(514)-398-7771
Fax:
(514)-398-7621
Email: info@eap.mcgill.ca
To report problems or otherwise comment on the structure of this site, send mail to the Webmaster